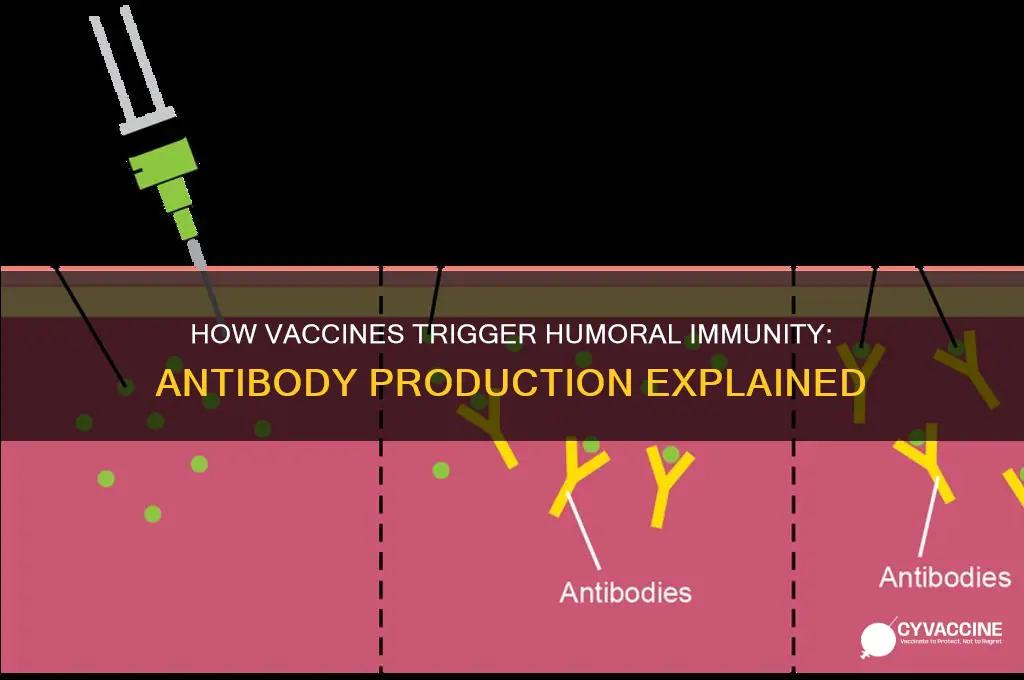
The humoral immune system, a critical component of the body's defense mechanism, plays a pivotal role in responding to vaccines by producing antibodies that neutralize pathogens. When a vaccine is administered, it introduces a harmless form of a pathogen, such as a weakened or inactivated virus, which triggers the immune system to recognize it as a threat. B lymphocytes, a type of white blood cell, are activated and differentiate into plasma cells that secrete specific antibodies tailored to bind to the pathogen's antigens. These antibodies circulate in the bloodstream and lymphatic system, ready to neutralize the actual pathogen if it ever invades the body. Additionally, memory B cells are generated, ensuring a faster and more robust antibody response upon future exposure to the same pathogen. This process not only provides immediate protection but also establishes long-term immunity, making the humoral immune response a cornerstone of vaccine efficacy.
Explore related products






What You'll Learn
![]()
Antigen presentation and B cell activation
The humoral immune response to vaccines is a complex process that relies heavily on antigen presentation and B cell activation. When a vaccine containing antigens (components of a pathogen) is administered, it is taken up by antigen-presenting cells (APCs), such as dendritic cells and macrophages. These APCs process the antigens into smaller peptides and display them on their surface bound to major histocompatibility complex class II (MHC-II) molecules. This antigen presentation is a critical step, as it allows the immune system to recognize foreign substances and initiate a targeted response. The APCs then migrate to lymphoid organs, such as lymph nodes or the spleen, where they encounter naïve B cells.
Naïve B cells express unique B cell receptors (BCRs) on their surface, which are specific to particular antigens. When a naïve B cell encounters an APC presenting a matching antigen, the BCR binds to the antigen-MHC-II complex. This binding event is the first signal required for B cell activation. However, B cell activation is not complete with just this signal. A second signal, known as co-stimulation, is necessary for full activation. Co-stimulatory molecules on the APC, such as CD80 and CD86, interact with their respective receptors (CD28) on the B cell, providing the additional signal needed for activation. This dual signaling ensures that B cells respond only to relevant antigens and prevents unnecessary immune reactions.
Once activated, B cells undergo proliferation and differentiation into either plasma cells or memory B cells. Plasma cells are the effector cells of the humoral immune response, responsible for producing and secreting antibodies specific to the antigen. These antibodies circulate in the bloodstream and lymphatic system, neutralizing pathogens or marking them for destruction by other immune cells. Memory B cells, on the other hand, persist long-term and provide a rapid and robust response upon secondary exposure to the same antigen, a principle that underlies the effectiveness of vaccines in conferring long-lasting immunity.
The role of T helper cells (Th cells) is also crucial in this process. Following antigen presentation by APCs, Th cells are activated and differentiate into Th2 cells, which secrete cytokines such as interleukin-4 (IL-4) and IL-5. These cytokines further stimulate B cell proliferation, class switching (allowing B cells to produce different classes of antibodies), and differentiation into plasma cells. Thus, the interaction between APCs, B cells, and Th cells is a highly coordinated process that ensures a robust and specific humoral immune response to vaccines.
In summary, antigen presentation and B cell activation are pivotal steps in the humoral immune response to vaccines. APCs process and present antigens to naïve B cells, which require both antigen recognition and co-stimulation for activation. Activated B cells then differentiate into plasma cells that produce antibodies and memory B cells that provide long-term immunity. The involvement of Th cells and their cytokines further amplifies and refines this response, ensuring that the immune system effectively neutralizes pathogens and establishes immunological memory. This intricate process highlights the elegance and efficiency of the humoral immune system in responding to vaccination.
Vaccination Injuries and Illnesses: Real Risks or Rare Occurrences?
You may want to see also
Explore related products






![]()
Production of vaccine-specific antibodies by plasma cells
The production of vaccine-specific antibodies by plasma cells is a cornerstone of the humoral immune response to vaccination. When a vaccine containing antigens (components of a pathogen) is introduced into the body, it triggers a cascade of immune reactions. Antigen-presenting cells (APCs), such as dendritic cells, engulf the vaccine antigens, process them, and present them on their surface via major histocompatibility complex (MHC) molecules. These APCs then migrate to lymph nodes, where they activate naive B cells that possess specific B-cell receptors (BCRs) complementary to the vaccine antigens. This activation marks the beginning of the humoral immune response.
Upon activation, naive B cells differentiate into plasmablasts, which are rapidly dividing cells committed to antibody production. Plasmablasts undergo further maturation and proliferation, eventually becoming plasma cells. Plasma cells are specialized, antibody-secreting cells that are the primary effectors of the humoral immune response. Each plasma cell is programmed to produce a single type of antibody, specific to the vaccine antigen that initiated the response. This specificity ensures that the antibodies generated are highly effective at recognizing and neutralizing the target pathogen.
The process of antibody production by plasma cells involves the synthesis of immunoglobulins (Igs), the Y-shaped proteins that function as antibodies. Plasma cells contain extensive endoplasmic reticulum and Golgi apparatus, which facilitate the mass production and secretion of antibodies. The antibodies produced are typically of the IgG class, which is the most abundant and versatile antibody isotype in the body. These IgG antibodies are released into the bloodstream and lymphatic system, where they circulate and bind to the vaccine antigens or the actual pathogen if future exposure occurs.
Vaccine-specific antibodies play multiple roles in immune protection. They can neutralize pathogens directly by blocking their ability to infect cells or by aggregating them for easier clearance. Antibodies also tag pathogens for destruction by other immune cells through processes like opsonization (coating pathogens to enhance phagocytosis) or activation of the complement system (a cascade of proteins that help eliminate pathogens). Additionally, some antibodies can prevent pathogens from adhering to host cells, thereby inhibiting infection.
The longevity of the humoral immune response is ensured by the formation of memory B cells, which are generated alongside plasma cells during the initial immune response. Memory B cells persist in the body for years or even decades, providing a rapid and robust antibody response upon re-exposure to the same pathogen. This is why vaccines often confer long-term immunity. Plasma cells, however, are short-lived, but their rapid production of antibodies during the initial response is critical for controlling the infection and establishing immunological memory. In summary, the production of vaccine-specific antibodies by plasma cells is a highly coordinated and efficient process that forms the basis of the humoral immune system's response to vaccines, ensuring both immediate and long-term protection against pathogens.
Vaccinated and Socializing: Understanding Your Contagiousness Post-Vaccination
You may want to see also
Explore related products




![]()
Memory B cell formation for long-term immunity
The humoral immune system plays a critical role in responding to vaccines by generating antibodies that neutralize pathogens. Central to this process is the formation of memory B cells, which provide long-term immunity against future infections. When a vaccine is administered, it introduces antigens that mimic a pathogen, triggering the immune system without causing disease. These antigens are taken up by antigen-presenting cells (APCs), which process and present them to naive B cells in lymphoid organs such as lymph nodes. Upon recognition of the antigen, naive B cells become activated and differentiate into antibody-secreting plasma cells and memory B cells. This initial activation is facilitated by T helper cells, which provide essential signals for B cell proliferation and class switching, enabling the production of high-affinity antibodies.
Memory B cell formation is a key outcome of this process, ensuring a rapid and robust response upon re-exposure to the same pathogen. Activated B cells undergo somatic hypermutation in germinal centers, where their antibody genes mutate to produce B cell receptors (BCRs) with higher affinity for the antigen. B cells with the highest affinity are selected for survival, while others undergo apoptosis. These high-affinity B cells then differentiate into either long-lived plasma cells, which continue to secrete antibodies, or memory B cells, which persist in the body for years or even decades. Memory B cells circulate in the bloodstream and lymphoid tissues, ready to mount a swift and effective response if the same antigen is encountered again.
The differentiation of memory B cells is influenced by various factors, including the type of antigen, the presence of cytokines, and interactions with T cells. Cytokines such as interleukin-21 (IL-21) and B cell-activating factor (BAFF) promote the survival and maintenance of memory B cells. Additionally, the strength and duration of the initial immune response impact the quality and quantity of memory B cells generated. Vaccines are designed to optimize this process by delivering antigens in a way that maximizes B cell activation and memory formation, often through adjuvants that enhance immune signaling.
Memory B cells provide long-term immunity by enabling a secondary immune response that is faster and more effective than the primary response. Upon re-exposure to the antigen, memory B cells rapidly proliferate and differentiate into plasma cells, producing a surge of high-affinity antibodies. This anamnestic response neutralizes the pathogen before it can cause disease, often preventing infection altogether. The persistence of memory B cells is maintained by their ability to self-renew and by the support of the bone marrow microenvironment, where some memory B cells reside.
In summary, memory B cell formation is a cornerstone of long-term immunity induced by vaccines. Through germinal center reactions, somatic hypermutation, and selection, high-affinity B cells differentiate into memory B cells that persist and provide rapid protection upon re-exposure to the antigen. This process is finely tuned by cytokines, T cell interactions, and the initial strength of the immune response. By harnessing the humoral immune system’s ability to generate memory B cells, vaccines establish durable immunity that safeguards individuals and populations from infectious diseases.
Polio Vaccine: Immunity and Protection
You may want to see also
Explore related products






![]()
Role of T helper cells in antibody class switching
The humoral immune response to vaccines is a complex process involving multiple components of the immune system, with T helper cells playing a pivotal role in orchestrating antibody production and class switching. When a vaccine is administered, it introduces antigens that are recognized by antigen-presenting cells (APCs), such as dendritic cells. These APCs process the antigens and present them to naive T helper cells (Th cells) in the lymph nodes. Upon recognition of the antigen-MHC complex, Th cells become activated and differentiate into effector Th cells, primarily Th2 cells, which are crucial for humoral immunity. Th2 cells secrete cytokines like IL-4, IL-5, and IL-13, which create a microenvironment conducive to B cell activation and differentiation.
T helper cells are essential for antibody class switching, a process that allows B cells to produce different classes of antibodies (e.g., IgG, IgA, IgE) with distinct effector functions. Class switching occurs in germinal centers of lymphoid tissues, where activated B cells undergo somatic hypermutation and isotype switching. Th2 cells provide critical signals to B cells through both cytokine secretion and direct cell-cell interaction via CD40-CD40L engagement. IL-4, in particular, is a key cytokine that induces the transcription of genes encoding IgG1 and IgE, promoting class switching away from the default IgM and IgG3 isotypes. Without Th cell help, B cells remain unable to undergo class switching, highlighting the indispensable role of Th cells in this process.
The interaction between T helper cells and B cells is tightly regulated to ensure appropriate antibody responses. Upon activation, B cells express surface receptors for cytokines produced by Th cells, such as IL-4 receptors, which further enhance their responsiveness to these signals. Additionally, the CD40-CD40L interaction provides a co-stimulatory signal that stabilizes the B cell-Th cell synapse and promotes sustained cytokine production. This prolonged interaction is vital for the initiation of class switching recombination (CSR), a DNA rearrangement process that replaces the constant region of the immunoglobulin gene, thereby altering the antibody class.
T helper cells also influence the affinity maturation of antibodies during class switching. In germinal centers, Th cells provide survival signals to B cells undergoing somatic hypermutation, allowing only those B cells producing high-affinity antibodies to persist. This selective process is mediated by cytokines and direct interactions, ensuring that the final antibody response is both effective and specific. Thus, Th cells not only drive class switching but also contribute to the overall quality of the humoral immune response.
In the context of vaccination, the role of T helper cells in antibody class switching is critical for generating long-lasting immunity. Vaccines often contain adjuvants that enhance Th cell activation, ensuring robust B cell responses and effective class switching. For example, aluminum salts, commonly used adjuvants, promote Th2-biased responses, favoring IgG1 production. Understanding the interplay between Th cells and B cells during class switching has significant implications for vaccine design, as it allows for the development of strategies to optimize antibody responses against specific pathogens.
In summary, T helper cells are central to antibody class switching in the humoral immune response to vaccines. Through cytokine secretion, co-stimulatory signals, and selective interactions with B cells, Th cells drive the production of diverse antibody classes and ensure the generation of high-affinity antibodies. Their role is not only instructive but also essential for the efficacy of vaccines, making them a critical focus in immunological research and vaccine development.
Medicare Coverage for Tetanus, Diphtheria, and Pertussis Vaccine
You may want to see also
Explore related products






![]()
Neutralization of pathogens by circulating antibodies post-vaccination
The humoral immune system plays a critical role in the body's defense against pathogens, particularly through the production and circulation of antibodies. Post-vaccination, the neutralization of pathogens by these circulating antibodies is a key mechanism that prevents infection and disease. When a vaccine is administered, it introduces a harmless form or component of the pathogen (such as a protein or inactivated virus) to the immune system. This triggers the activation of B cells, a type of white blood cell, which differentiate into plasma cells. These plasma cells then secrete antibodies specific to the pathogen's antigens, which are unique molecular markers on the pathogen's surface. These antibodies are released into the bloodstream and lymphatic system, where they circulate and remain on standby to combat future encounters with the actual pathogen.
Circulating antibodies neutralize pathogens primarily by binding to their specific antigens, a process that can directly block the pathogen's ability to infect host cells. For example, in the case of viral pathogens, antibodies can attach to viral surface proteins, preventing the virus from attaching to and entering host cells. This is known as viral neutralization. Similarly, for bacterial pathogens, antibodies can bind to toxins or surface structures, rendering them ineffective or marking them for destruction by other immune cells. This binding not only inhibits the pathogen's ability to cause harm but also facilitates its clearance from the body. The specificity of this interaction ensures that the immune response is targeted and effective, minimizing damage to healthy tissues.
Another important aspect of antibody-mediated neutralization is the activation of the complement system, a cascade of proteins that enhances the immune response. When antibodies bind to pathogens, they can trigger the complement pathway, leading to the formation of a membrane attack complex that lyses (destroys) the pathogen's cell membrane. Additionally, antibodies can opsonize pathogens, meaning they coat the pathogen's surface, making it easier for phagocytic cells like macrophages and neutrophils to engulf and destroy them. This dual action of direct neutralization and immune system recruitment significantly enhances the body's ability to eliminate pathogens before they can establish an infection.
The longevity of circulating antibodies post-vaccination is a critical factor in maintaining immunity. Memory B cells, which are generated during the initial immune response, persist in the body for years or even decades. Upon re-exposure to the same pathogen, these memory B cells rapidly proliferate and differentiate into plasma cells, producing a robust and swift antibody response. This secondary response is typically faster and more effective than the primary response, often preventing the pathogen from causing symptomatic disease. Vaccines are designed to maximize this long-term immunity by inducing the formation of a robust pool of memory B cells and circulating antibodies.
In summary, the neutralization of pathogens by circulating antibodies post-vaccination is a multifaceted process that involves direct binding to antigens, activation of the complement system, opsonization, and the rapid secondary response mediated by memory B cells. This mechanism is central to the humoral immune system's ability to provide durable protection against infectious diseases. Understanding these processes underscores the importance of vaccination in harnessing the immune system's power to prevent illness and save lives.
Vaccination Requirements for Entry into Kenya
You may want to see also
Frequently asked questions
The humoral immune system responds to vaccines by producing antibodies, primarily through the activation of B cells. When a vaccine is administered, it introduces antigens (components of the pathogen) that stimulate B cells to differentiate into plasma cells. These plasma cells then secrete antibodies specific to the antigen, which can neutralize pathogens or mark them for destruction by other immune cells.
During a humoral immune response to vaccines, the body initially produces IgM antibodies, which are the first line of defense. Over time, B cells undergo class switching and affinity maturation, leading to the production of higher-affinity IgG antibodies. IgG antibodies are more effective at neutralizing pathogens and provide long-term immunity, contributing to immunological memory.
The humoral immune system contributes to long-term immunity by generating memory B cells during the initial immune response to a vaccine. These memory B cells persist in the body and can rapidly produce high levels of specific antibodies upon re-exposure to the same pathogen. This quick response prevents infection and ensures lasting protection against the disease.